Способы размножения грибов
Грибы размножаются вегетативным, бесполым и половым путями.
Вегетативное размножение происходит без образования специализированных органов – любая часть мицелия дает начало новому организму. Вегетативное размножение применяют при поддержании культуры на искусственных питательных средах.
Одной из форм вегетативного размножения является образование хламидоспор – клеток, покрытых толстой оболочкой и способных переносить длительные периоды голодания, высыхания и другие неблагоприятные условия среды.
Вегетативным размножением является также почкование клеток и фрагментация гиф на цилиндрические или овальные клетки – оидии, образованию которых предшествует появление частых перегородок в гифах. В благоприятных условиях оидии прорастают в новый мицелий.
Бесполое размножение грибов происходит при помощи особых спор, появлению которых не предшествует предварительное слияние клеток и объединение ядер. У одних грибов споры развиваются эндогенно в специальных вместилищах, спорангиях, и называются спорангиоспорами. У других – споры формируются экзогенно на специализированных гифах мицелия, конидиеносцах, и называются конидиями. Для зигомицетов характерно бесполое размножение при помощи спорангиоспор; для аскомицетов, базидиомицетов, дейтеромицетов и некоторых низших грибов – при помощи конидий.
В отличие от спорангиоспор и спорангиеносцев конидии и конидиеносцы разнообразны по строению. Конидиеносцы различаются по типу ветвления (мутовчатое, моноподиальное, дихотомическое, симподиальное), имеют различные вздутия, стеригмы, фиалиды. Конидиеносцы многих грибов бывают объединены в коремии, ложа, и пикниды.
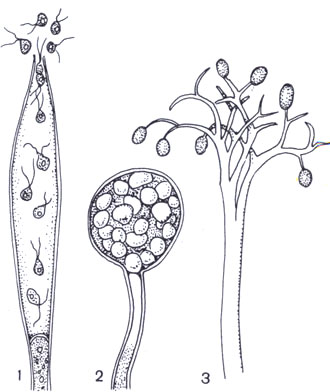
Коремии представляют собой пучок тесно сближенных конидиеносцев (рис.1). Подобные образования характерны для некоторых пенициллов. У дейтеромицетов конидиеносцы часто располагаются тесным слоем на специальной поверхности, состоящей из плоского сплетения гиф, образуя ложе. Наиболее сложной формой агрегации конидиеносцев являются полузамкнутые структуры – пикниды. Они содержат большое количество конидиеносцев, хорошо защищенных оболочкой.
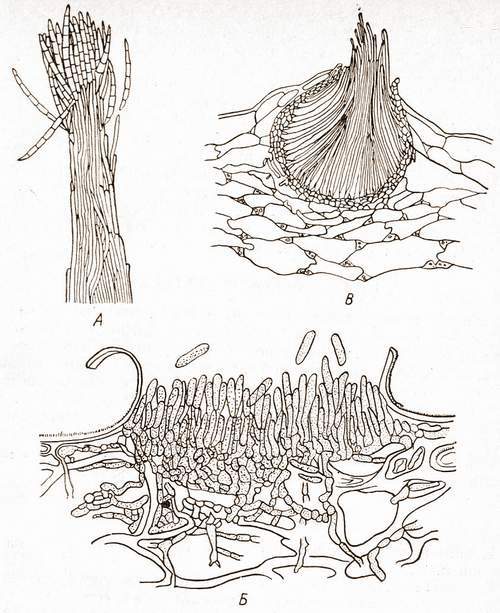
Рис.1. Конидиеносцы грибов :
А – коремия Sphaerostilbe ; Б – ложе Gloeosporium ; В – пикнида Septoria
Конидии грибов так же, как конидиеносцы, очень разнообразны по своему строению. Они бывают одноклеточные и с перегородками; различаются по форме (шаровидные, эллипсовидные, звездчатые, спирально закрученные и др.) и цвету (темные, буровато-коричневые, светлые).
Грибы образуют очень большое количество спор бесполого размножения. Известно, например, что трутовый гриб Ganoderma applanatum в течение 6 месяцев способен ежедневно высвобождать до 30х10 6 спор; а колония пеницилла диаметром 2,5 см может образовать 400х10 6 конидий. Распространение спор грибов, вызывающих повреждения материалов, обычно происходит при помощи ветра, дождя, насекомых, животных и человека.
Половое размножение установлено для всех групп грибов, кроме дейтеромицетов, у которых половые стадии отсутствуют. Оно происходит при помощи специальных спор. Образованию их обязательно предшествует слияние клеток, часто специализированных, и последующее объединение ядер.
Половой процесс состоит из трех стадий: плазмогамии, кариогамии и мейоза. В первую фазу происходит объединение протоплазменного содержимого двух клеток. Во вторую – слияние ядер и образование диплоидного ядра с двойным набором хромосом. Третья стадия характеризуется редукционным делением ядра с восстановлением гаплоидного набора хромосом. Конечным результатом полового воспроизведения у всех грибов является образование специальных спор, служащих для размножения. У низших грибов это споры зародышевого спорангия, у высших – аскоспоры и базидиоспоры.
Споры, возникшие половым путем, обеспечивают главным образом сохранение вида в течение периода, неблагоприятного для вегетации, а также способствуют генетическому разнообразию популяции. Споры бесполого размножения служат для быстрой колонизации субстрата. Заражение промышленных материалов происходит в основном с помощью спор бесполого размножения.
Парасексуальный процесс . Некоторые грибы не имеют истинного полового процесса. Обмен генетической информацией у них происходит в результате парасексуального цикла. Он обнаружен у многих дейтеромицетов и базидиомицетов главным образом в лабораторных условиях. В результате этого процесса плазмогамия, кариогамия и гаплоидизация хромосом происходят не в специализированных половых структурах, или в определенной стадии жизненного цикла, а в вегетативной гифе мицелия. Парасексуальный цикл заключается в слиянии генетически различных ядер в любой вегетативной гифе гетерокариотического мицелия и последующей рекомбинации участков хромосом при митозе.
Парасексуальный цикл имеет большое значение у несовершенных грибов, для которых характерно отсутствие полового процесса. Например, для грибов рода Verticillium он является источником возникновения изменчивости. У некоторых грибов парасексуальный цикл происходит и при наличии полового процесса.
Споры грибов

Митоспоры и мейоспоры выполняют различные функции в жизни грибов. Митоспоры предназначены, в основном, для массового расселения грибов в вегетативный период, без рекомбинации наследственных качеств. Мейоспоры являются спорами полового размножения. Их появление связано с прорастанием зиготы – клетки, образованной при половом процессе из двух родительских клеток. В процессе прорастания зиготы и формировании спор полового размножения осуществляется рекомбинация (перераспределение) признаков родительских организмов, и увеличивается разнообразие потомков.
У многих видов грибов в жизненном цикле происходит образование как митоспор, так и мейоспор. К примеру, у многих сумчатых грибов – возбудителей спорыньи, мучнистой росы растений, парши яблок и др. митоспоры формируются на протяжении всего периода вегетации и выполняют функцию повторного заражения растений, а мейоспоры образуются после зимовки и провоцируют первичное заражение растений.
Формирование
Формирование митоспор у большинства форм грибов происходит внутри особых клеток – спорангиев либо наружно на веточках воздушного мицелия. Первые виды спор называются спорангиоспорами, вторые – конидиями. Спорангиоспоры бывают неподвижными и переносяться пасивно, могут иметь жгутики и с их помощью активно передвигаться в водной среде. Подвижные споры носять название зооспоры. Размножение зооспорами осуществляется только при наличии капельно-жидкой среды.
Спорангиоспоры, как и зооспоры, служат для широкого расселения грибов. Они содержат небольшое количество питательных веществ и не способны длительно сохранять свою жизнеспособность. В одном спорангии может формироваться до нескольких десятков тысяч спор. Увеличение выработки спорангиоспор достигается за счет дополнительных ответвлений спорангиеносцев и увеличения количества спорангиев. Поэтому для образования крупного спорангия с множеством спорангиоспор нужен более длительный срок, чем для массового формирования конидий. Этим объясняется то, что у большинства грибов распространено бесполое размножение с помощью конидий.  Конидии образуются у высших грибов с клеточным мицелием на гифах воздушного мицелия – конидиеносцах. Они обычно очень мелкие (от 1 до 10-15 мкм) и формируются в огромных количествах. В отличие от спорангиоспор, конидии могут длительно сохранять жизнеспособность в неблагоприятных условиях, не прорастая. Конидии по мере созревания высвобождаются как активно, так и пассивно. У них в процессе эволюции сформировались множественные адаптации, облегчающие их распространение. У некоторых конидий сухая поверхность, благодаря чему они легко переносятся ветром; другие покрыты слизью и хорошо переносяться насекомыми или водой, т.д.
Конидии образуются у высших грибов с клеточным мицелием на гифах воздушного мицелия – конидиеносцах. Они обычно очень мелкие (от 1 до 10-15 мкм) и формируются в огромных количествах. В отличие от спорангиоспор, конидии могут длительно сохранять жизнеспособность в неблагоприятных условиях, не прорастая. Конидии по мере созревания высвобождаются как активно, так и пассивно. У них в процессе эволюции сформировались множественные адаптации, облегчающие их распространение. У некоторых конидий сухая поверхность, благодаря чему они легко переносятся ветром; другие покрыты слизью и хорошо переносяться насекомыми или водой, т.д.
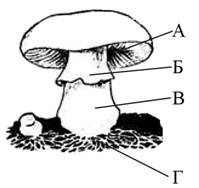
Задача 8749 Какой буквой на рисунке обозначена часть
УСЛОВИЕ:
Какой буквой на рисунке обозначена часть шляпочного гриба, где происходит образование спор?
РЕШЕНИЕ:
ответ а
тк споры образуются в спорангиях-в трубочках или на пластинках шляпки
![]()
ОТВЕТ:
Добавил OlyaRusinova , просмотры: O 1964 ? 28.04.2016. биология класс не задан класс
Решения пользователей
Написать комментарий
На шахматной доске mn клеток.
Первую ладью можно поставить на любое из mn мест.
Ладья ходит по горизонтали и вертикали.
Вычеркиваем горизонталь и вертикаль на которых она стоит.
Получаем (m-1)*(n-1) клеток, на которые можно поставить вторую ладью.
(m-[red]2[/red])*(n-[red]2)[/red] клеток, на которые можно поставить [red]третью[/red] ладью
.
(m-([green]k-1[/green]))*(n-([green]k-1[/green])) клеток, на которые можно поставить [green]k-ую[/green] ладью
По правилу умножения эти выборы надо умножить и разделить на перестановку из k
элементов
Выбираем шесть человек из десяти.
Это можно сделать C^(6)_(10)=10!/(6!*4!)=(7*8*9*10)/24=210 способов.
Т. е. имеем 210 вариантов списка состава участников.
В первый день можно взять один состав из 210, во второй день — один из оставшихся 209, в третий — один из оставшихся 208
Выбор в течение трех дней это выбор тройки ( состав первого дня; состав второго дня; состав третьего дня) можно осуществить
210*209*208= считайте
![]()
5+ 7 +3=15 фруктов
Но мы не должны учитывать перестановки когда объекты одного типа меняются местами.
Поэтому нужно поделить 15! на (5!*7!*3!)
О т в е т. (15!)/((5!*7!*3!)=(8*9*10*11*12*13*14*15)/(1*2*3*4*5*2*3)=11*12*13*14*15
умножьте и получите ответ
Лекция № 14. Размножение организмов
Размножение — свойство живых организмов воспроизводить себе подобных. Существуют два основных способа размножения — бесполое и половое.
Бесполое размножение
Бесполое размножение осуществляется при участии лишь одной родительской особи и происходит без образования гамет. Дочернее поколение у одних видов возникает из одной или группы клеток материнского организма, у других видов — в специализированных органах. Различают следующие способы бесполого размножения: деление, почкование, фрагментация, полиэмбриония, спорообразование, вегетативное размножение.

Деление — способ бесполого размножения, характерный для одноклеточных организмов, при котором материнская особь делится на две или большее количество дочерних клеток. Можно выделить: а) простое бинарное деление (прокариоты), б) митотическое бинарное деление (простейшие, одноклеточные водоросли), в) множественное деление, или шизогонию (малярийный плазмодий, трипаносомы). Во время деления парамеции (1) микронуклеус делится митозом, макронуклеус — амитозом. Во время шизогонии (2) сперва многократно митозом делится ядро, затем каждое из дочерних ядер окружается цитоплазмой, и формируются несколько самостоятельных организмов.

Почкование — способ бесполого размножения, при котором новые особи образуются в виде выростов на теле родительской особи (3). Дочерние особи могут отделяться от материнской и переходить к самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом случае колонии (коралловые полипы).
Фрагментация (4) — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды, спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации.
Полиэмбриония — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается эмбрион (монозиготные близнецы).
Вегетативное размножение — способ бесполого размножения, при котором новые особи образуются или из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.), специально предназначенных для этой формы размножения. Вегетативное размножение характерно для многих групп растений, используется в садоводстве, огородничестве, селекции растений (искусственное вегетативное размножение).
| Вегетативный орган | Способ вегетативного размножения | Примеры |
|---|---|---|
| Корень | Корневые черенки | Шиповник, малина, осина, ива, одуванчик |
| Корневые отпрыски | Вишня, слива, осот, бодяк, сирень | |
| Надземные части побегов | Деление кустов | Флокс, маргаритка, примула, ревень |
| Стеблевые черенки | Виноград, смородина, крыжовник | |
| Отводки | Крыжовник, виноград, черемуха | |
| Подземные части побегов | Корневище | Спаржа, бамбук, ирис, ландыш |
| Клубень | Картофель, седмичник, топинамбур | |
| Луковица | Лук, чеснок, тюльпан, гиацинт | |
| Клубнелуковица | Гладиолус, крокус | |
| Лист | Листовые черенки | Бегония, глоксиния, колеус |
Спорообразование (6) — размножение посредством спор. Споры — специализированные клетки, у большинства видов образуются в особых органах — спорангиях. У высших растений образованию спор предшествует мейоз.
Клонирование — комплекс методов, используемых человеком для получения генетически идентичных копий клеток или особей. Клон — совокупность клеток или особей, произошедших от общего предка путем бесполого размножения. В основе получения клона лежит митоз (у бактерий — простое деление).
Половое размножение

Половое размножение осуществляется при участии двух родительских особей (мужской и женской), у которых в особых органах образуются специализированные клетки — гаметы. Процесс формирования гамет называется гаметогенезом, основным этапом гаметогенеза является мейоз. Дочернее поколение развивается из зиготы — клетки, образовавшейся в результате слияния мужской и женской гамет. Процесс слияния мужской и женской гамет называется оплодотворением. Обязательным следствием полового размножения является перекомбинация генетического материала у дочернего поколения.
В зависимости от особенностей строения гамет, можно выделить следующие формы полового размножения: изогамию, гетерогамию и овогамию.
Изогамия (1) — форма полового размножения, при которой гаметы (условно женские и условно мужские) являются подвижными и имеют одинаковые морфологию и размеры.
Гетерогамия (2) — форма полового размножения, при которой женские и мужские гаметы являются подвижными, но женские — крупнее мужских и менее подвижны.
Овогамия (3) — форма полового размножения, при которой женские гаметы неподвижные и более крупные, чем мужские гаметы. В этом случае женские гаметы называются яйцеклетками, мужские гаметы, если имеют жгутики, — сперматозоидами, если не имеют, — спермиями.
Овогамия характерна для большинства видов животных и растений. Изогамия и гетерогамия встречаются у некоторых примитивных организмов (водоросли). Кроме вышеперечисленных, у некоторых водорослей и грибов имеются формы размножения, при которых половые клетки не образуются: хологамия и конъюгация. При хологамии происходит слияние друг с другом одноклеточных гаплоидных организмов, которые в данном случае выступают в роли гамет. Образовавшаяся диплоидная зигота затем делится мейозом с образованием четырех гаплоидных организмов. При конъюгации (4) происходит слияние содержимого отдельных гаплоидных клеток нитевидных талломов. По специально образующимся каналам содержимое одной клетки перетекает в другую, образуется диплоидная зигота, которая обычно после периода покоя также делится мейозом.
Перейти к лекции №13 «Способы деления эукариотических клеток: митоз, мейоз, амитоз»
Перейти к лекции №15 «Половое размножение у покрытосеменных растений»
Смотреть оглавление (лекции №1-25)
Как происходит образование спор у грибов
Что ты хочешь узнать?
Ответ

Споры грибов предназначены для размножения. Они представляют собой одну либо несколько клеток, имеющих микроскопические размеры – от 1 до 100 мк. Споры большинства видов грибов содержат крайне мало питательных веществ, редкие из них выживают и, попав в благоприятные условия, прорастают, давая начало мицелию молодого гриба. Но гибель большинства спор компенсируется формированием их огромного множества. К примеру, возбудитель стеблевой ржавчины злаковых растений образует весной и в июне на одном кусте барбариса до 10 млрд. спор, плодовое тело трутовика стандартных размеров дает до 30 млрд. спор, шампиньоны образуют примерно 40 млрд. спор в час.
Общепринята классификация спор по происхождению и выполняемой функции в жизни грибов на две обширные группы:
а) споры бесполого размножения (митоспоры);
б) споры полового размножения (мейоспоры).
При образовании митоспор происходит митотическое деление клеточного ядра с сохранением числа хромосом; мейоспор – мейоз, характеризующийся уменьшением количества хромосом в два раза.
У большой группы грибов митоспор в цикле развития нет, как у многих шляпочных грибов, трутовиков, гастеромицетов, некоторых видов дискомицетов (сморчков, строчков). В таких случаях мейоспоры берут на себя функции митоспор и служат для массового расселения грибов.
Спорангиоспоры, как и зооспоры, служат для широкого расселения грибов. Они содержат небольшое количество питательных веществ и не способны длительно сохранять свою жизнеспособность. В одном спорангии может формироваться до нескольких десятков тысяч спор. Увеличение выработки спорангиоспор достигается за счет дополнительных ответвлений спорангиеносцев и увеличения количества спорангиев. Поэтому для образования крупного спорангия с множеством спорангиоспор нужен более длительный срок, чем для массового формирования конидий. Этим объясняется то, что у большинства грибов распространено бесполое размножение с помощью конидий.
Конидии образуются у высших грибов с клеточным мицелием на гифах воздушного мицелия – конидиеносцах. Они обычно очень мелкие (от 1 до 10-15 мкм) и формируются в огромных количествах. В отличие от спорангиоспор, конидии могут длительно сохранять жизнеспособность в неблагоприятных условиях, не прорастая. Конидии по мере созревания высвобождаются как активно, так и пассивно. У них в процессе эволюции сформировались множественные адаптации, облегчающие их распространение. У некоторых конидий сухая поверхность, благодаря чему они легко переносятся ветром; другие покрыты слизью и хорошо переносяться насекомыми или водой, т.д.
Мейоспоры, как и митоспоры, у грибов чрезвычайно разнообразны. Они образуются внутри либо на поверхности специальных клеток, которые называются, соответственно, сумками и базидиями. Сумки находятся в плодовых телах грибов и хорошо защищены от воздействия неблагоприятных факторов внешней среды. У грибов из группы аскомицетов образуются аскоспоры, у базидиомицетов – базидиоспоры, имеющих разное строение.
Где происходит образование спор у грибов




Размножение грибов
Размножение грибов происходит при помощи спор, а также кусочков мицелия, склероциев и других структур. Любой элемент, который может дать начало новому мицелию, часто называют диаспорой или пропагулой. По происхождению диаспоры могут быть специализированными, т. е. образующимися специально для размножения (споры), или неспециализированными (например, фрагменты мицелия).
Размножение неспециализированными диаспорами получило название вегетативного, так как оно происходит за счет участков или структур вегетативного тела грибов. Мицелиальные грибы чаще размножаются кусочками мицелия. Этот способ обычно используют для размножения многих грибов, например шампиньонов, вешенки, летнего опенка и др., в культуре.
Споры грибов служат для размножения. Они состоят из одной или нескольких клеток и имеют микроскопические размеры — от 1-2 до 100 мк и более. За относительно немногими исключениями, споры содержат мало запасных питательных веществ, только немногие выживают и, попав в подходящие условия, прорастают, давая начало новому мицелию. Однако гибель компенсируется образованием огромного количества спор. Так, возбудитель стеблевой ржавчины злаков образует весной и в начале лета на одном кусте барбариса до 64 • 1012 спор, плодовое тело трутовика средних размеров до 25-30 млрд., а шампиньоны около 40 млн. спор в час.
По происхождению и роли в жизни грибов споры делятся на две большие группы: споры бесполого размножения, или митоспоры, и споры полового размножения, или мейоспоры. Образованию митоспор предшествует митотическое деление клеточного ядра, т. е. деление, не сопровождающееся сокращением числа хромосом. В противоположность этому образованию мейоспор предшествует мейоз — деление ядра клетки, сопровождающееся сокращением числа хромосом вдвое.
Митоспоры и мейоспоры различаются не только происхождением, но функциями. Митоспоры служат преимущественно для массового расселения грибов в период вегетации, без рекомбинации наследственных признаков. При бесполом размножении в принципе происходит образование точных копий, идентичных родителям, а изменчивость возникает в основном за счет мутаций — наследственных изменений в генах. Однако в природе мы встречаемся обычно с большой изменчивостью грибов, размножающихся митоспорами. Это связано с образованием огромных их количеств, при которых даже при низкой частоте мутаций те из них, которые благоприятны для жизни гриба в существующих условиях, сохраняются и накапливаются, создавая большое разнообразие в его природных популяциях.
Мейоспоры — споры полового размножения. Они образуются при прорастании зиготы — клетки, возникшей при половом процессе из двух родительских клеток. При прорастании зиготы и образовании спор полового размножения происходит перераспределение (рекомбинация) признаков родительских форм и увеличивается разнообразие.
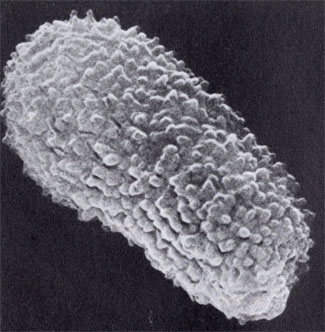
Так выглядит спора гриба под электронным сканирующим микроскопом, увеличенная в 20 тыс. раз
В жизненном цикле многих грибов образуются и митоспоры, и мейоспоры. Например, у многих сумчатых грибов — возбудителей мучнистой росы растений, парши яблок и груш, спорыньи и др. митоспоры образуются в течение периода вегетации и служат для повторного заражения растений, а мейоспоры — после перезимовки и вызывают первичное заражение растений. Однако у многочисленных грибов митоспоры в цикле развития отсутствуют. Нет их у большинства шляпочных грибов, гастеромицетов, трутовиков, многих дискомицетов, например сморчков и строчков, и др. В этих случаях мейоспоры принимают на себя функции митоспор — служат для массового расселения грибов. Известна и большая группа грибов, называемых дейтеромицетами (около 30 000 видов), у которых, наоборот, полностью утрачена способность к половому размножению и образуются только митоспоры.
Митоспоры образуются у грибов внутри специальных клеток — спорангиев или на веточках воздушного мицелия. Первые носят название спорангиоспоры и образуются путем раскалывания протопласта спорангия с многочисленными ядрами на одноядерные участки, каждый из которых затем превращается в спору. Митоспоры, образующиеся экзогенно (наружно) на веточках воздушного мицелия, называются конидиями.
Спорангиоспоры могут быть неподвижными и переноситься пассивно или имеют жгутики и активно движутся. Подвижные споры со жгутиками называют зооспорами. Зооспора представляет собой комочек протоплазмы, лишенный жесткой клеточной стенки и снабженный одним или двумя жгутиками. Они существуют совсем недолго, от нескольких минут до нескольких часов, и сохраняют жизнеспособность только в воде. Поэтому размножение зооспорами может происходить только в присутствии капельно-жидкой среды и дает преимущества только при обитании в воде. Проплавав определенное время, зооспора оседает на подходящий субстрат и прорастает в новое вегетативное тело. Зооспоры способны активно перемещаться лишь на небольшие расстояния (до 10 см) и с относительно небольшой скоростью, поэтому в их распространении большое значение имеет и пассивный перенос их токами воды. Однако для многих грибов — паразитов растений, размножающихся зооспорами, способность последних к активному движению даже на небольшие расстояния очень важна для заражения хозяина. Их зооспоры обладают положительным хемотаксисом — способностью активно двигаться по направлению к выделениям корней хозяев, содержащим аминокислоты, сахара и другие вещества. В результате такого движения зооспоры скапливаются у корней хозяина и вызывают их заражение. Особенно часто это наблюдается у различных паразитов на корнях растений, например у питиумов, ольпидиума капустного — возбудителя «черной ножки» рассады капусты и др.
Размножение зооспорами мы встречаем у всех групп первично водных грибов. У наземных грибов способность к их образованию постепенно утрачивается.
Типы бесполового размножения у грибов: 1 — зооспорангий с зооспорами; 2 — спорангий со спорангиоспорами; 3 — конидиеносец с конидиями
Одна из наиболее интересных в биологическом отношении групп грибов — пероно-споровые. В этой группе, происшедшей от водных грибов, только немногие ее представители связаны сейчас с водной средой, а большинство других — наземные организмы, преимущественно паразиты цветковых растений. Поэтому в ней хорошо прослеживаются те изменения, которые произошли у грибов при выходе их на сушу. У многих наземных грибов из пероноспоровых сохранилось еще размножение зооспорами, но у них возникли и многочисленные приспособления для распространения в воздушной среде. Так, зооспорангий у них не прорастает на месте его образования, как это происходит у водных грибов, а отделяется от спорангиеносца и переносится целиком, как конидия, ветром или каплями дождя. Зооспоры выходят из такого зооспорангия только в присутствии капельно-жидкой воды и при пониженной температуре воздуха, а при повышенной температуре в сухую погоду зооспорангий прорастает непосредственно ростковой трубкой. Это наблюдается, например, у фитофторы — возбудителя фитофтороза у картофеля и томатов.
У мукоровых грибов, образующих неподвижные спорангиоспоры, число их в одном спорангии колеблется от одной до 70 000 тысяч в больших спорангиях, диаметром до 1,5 мм, например у фикомицеса Блексли. Увеличение продукции спорангиоспор достигается у них также ветвлением спорангиеносцев и соответственно увеличением числа спорангиев. Спорангиоспоры, как и зооспоры, служат только для расселения грибов, они содержат мало питательных веществ и не способны сохранять жизнеспособность в течение длительного времени. Кроме того, для образования большого спорангия с многочисленными спорангиоспорами нужно значительно больше времени, чем для массового образования конидий. Поэтому у большинства грибов получило распространение бесполое размножение при помощи конидий.
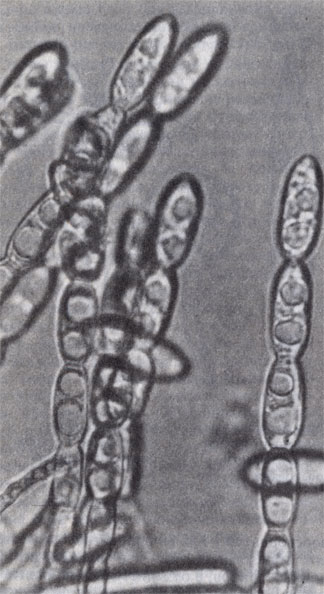
Конидиальное спороношение мучнисторосяного гриба
Образование конидий характерно для грибов с клеточным мицелием, так называемых высших грибов. Конидии образуются на гифах воздушного мицелия, обычно на специализированных веточках — конидиеносцах. Они могут быть очень мелкими, размером от 1 до 10-15 мкм, и формироваться в огромных количествах. Так, одна колония пеницилла (зеленой плесени) диаметром не более 3 см образует около 4-Ю8 конидий. У других грибов конидии размером до 50-100 мк и более. Такие конидии часто многоклеточные, толстостенные, содержат в клеточных стенках темные пигменты — меланины. В отличие от спорангиоспор конидии грибов могут часто длительно сохранять жизнеспособность при неблагоприятных условиях, не прорастая.
Конидиеносцы образуются непосредственно на воздушном мицелии поодиночке, как его веточки, или собраны в большие группы. У цератоцистиса вязового — возбудителя голландской болезни вязов конидиеносцы срастаются в длинный столбик, на вершине которого образуются конидии. Нектрия киноварно-красная, часто встречающаяся на ветках различных лиственных деревьев и кустарников, образует розовые подушечки плотно собранных конидиеносцев. У некоторых паразитов растений конидиеносцы собраны плотным слоем под кутикулой или эпидермисом пораженных листьев. Наконец, конидиеносцы могут развиваться внутри специальных, обычно шаровидных или грушевидных вместилищ, имеющих на вершине отверстие, через которое освобождаются конидии. Они обеспечивают не только массовую продукцию конидий, но и защищают развивающиеся конидиеносцы и конидии от неблагоприятных воздействий внешней среды.
Освобождение конидий происходит, как правило, пассивно, хотя у некоторых грибов — нигроспоры, дейгтониеллы и др. — наблюдается активное их отбрасывание. У конидий возникли и многочисленные адаптации, облегчающие их распространение. Некоторые конидии имеют сухую, плохо смачивающуюся поверхность и легко переносятся токами воздуха, другие погружены в слизь и переносятся водой или насекомыми. У грибов, вторично перешедших к обитанию в водной среде, конидии часто приобретают причудливую форму — тетрарадиальную, извитую, спиральную и т. п. Такая форма обеспечивает длительное парение конидий в толще воды, они не тонут и переносятся токами воды на большие расстояния.
Мейоспоры грибов, как и митоспоры, тоже крайне разнообразны. У грибов, образующих спорангиоспоры, споры полового размножения также образуются в специальных спорангиях — зародышевых. Но у большей части известных сейчас грибов (более чем у 60 000 видов) мейоспоры образуются внутри или на поверхности специальных клеток, называемых соответственно сумками и базидиями.
Рассматривая под микроскопом тонкий срез сморчка, строчка или чашечку пецицы, можно увидеть довольно правильный слой, состоящий из удлиненных клеток и расположенных между ними более тонких, нитевидных гиф. Если плодовое тело было зрелым, в удлиненных цилиндрических клетках будет видно по 8 спор, расположенных в один ряд. Эти цилиндрические клетки и называют сумками или асками, а образующиеся в них мейоспоры — сумкоспорами или аскоспорами. Образование сумок характерно для очень большой группы грибов — класса аскомицетов, или сумчатых. Однако далеко не у всех из них образуются такие крупные и хорошо развитые плодовые тела, как у сморчков и строчков. У наиболее примитивных аскомицетов сумки появляются прямо на мицелии, а аскоспоры освобождаются из них пассивно после разрыва или расплывания оболочки сумки.
В процессе эволюции у аскомицетов возникли плодовые тела, сначала очень мелкие и примитивные, с беспорядочно разбросанными внутри них сумками, а затем все более и более совершенные, в которых сумки располагались правильным слоем — гимением. В плодовых телах сумки хорошо защищены, особенно на ранних стадиях их развития, от влияния неблагоприятных факторов среды — высушивания, излишней инсоляции, колебания температуры и т. п. В процессе эволюции аскомицеты стали активно выбрасывать аскоспоры из сумок. Аскоспоры в сумке окружены не использованной на их формирование цитоплазмой. К моменту созревания аскоспор в ней происходит превращение гликогена в сахар, тургорное давление в сумке резко возрастает, оболочка ее разрывается и аскоспоры с силой выбрасываются на расстояние от десятых долей миллиметра до нескольких десятков сантиметров.
У аскомицетов известно несколько типов плодовых тел. Наиболее примитивные из них — клейстотеции. Они имеют обычно шаровидную или сплюснутую форму, полностью замкнуты, и размеры их невелики, 100-500 мкм в диаметре, реже крупнее. В культуре на питательных средах или в природе их часто можно заметить невооруженным глазом в виде маленьких, различной окраски шариков — белых, желтых, оранжевых, бурых или черных.
Более совершенный тип плодового тела — перитеций. Это полузамкнутое плодовое тело, большей частью кувшинообразное или округлое, с узким выводным отверстием на вершине, на дне расположены пучком или слоем сумки и стерильные гифы — парафизы. Аскоспоры выбрасываются из перитеция обычно активно. Этот процесс детально описан у многих аскомицетов. У сордарии, например, обильно развивающейся на навозе травоядных животных, сумки, удлиняясь за счет растяжения или роста, поочередно достигают отверстия перитеция и выбрасывают аскоспоры на расстояние до 20 см. У некоторых грибов, имеющих перитеции с очень длинной шейкой, например у эндотии, вызывающей серьезное заболевание каштанов, сумки имеют быстро растворяющуюся ножку и после созревания свободно лежат в слизи в полости перитеция. При набухании сумок в перитеции создается давление, и они поочередно выталкиваются к отверстию, выбрасывая аскоспоры. За час из перитеция выбрасывается около 14 000 аскоспор.
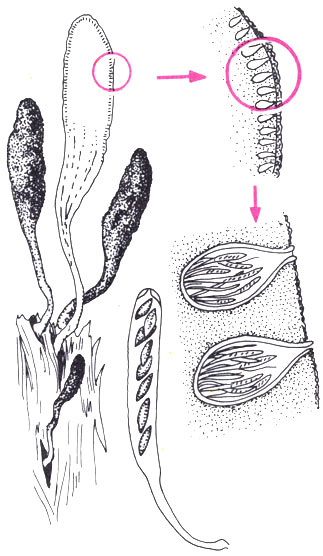
У сумчатого гриба ксилярии перитеции образуются на стромах и в них содержится масса сумок с аскоспорами
Наиболее совершенный тип плодового тела у аскомицетов — апотеций. Это широкораскрытые при созревании плодовые тела, большей частью блюдцевидные, чашевидные, бокаловидные или дисковидные. Их верхняя сторона — диск — выстлана гимением, состоящим из сумок и парафиз. Аскоспоры из апотециев выбрасываются активно. Расположение сумок широко открытым гимением дает возможность одновременного выбрасывания аскоспор из многих сумок в виде взрыва или залпа. При прикосновении к крупным апотециям пецицы или других дискомицетов часто можно увидеть над ними легкое облачко выброшенных аскоспор и даже слышать характерный щелчок. Аскоспоры выбрасываются из апотециев на расстояние от 0,5 до 20, а у некоторых видов до 60 см. Иногда строение апотециев отличается от типичного, чашевидного. Они бывают булавовидные у геоглоссума, часто встречающегося в хвойно-широколиственных лесах, имеют вид лопаточки у спатулярии, шляпки с закрученным краем на ножке у кудонии, обычной в наших еловых или смешанных лесах. У сморчков, строчков и сморчковой шапочки апотеций дифференцированы на шляпку и ножку, а клубневидные апотеции трюфелей вторично замкнуты в связи с обитанием под землей. Апотеции — самые крупные плодовые тела у аскомицетов, их размеры — от долей миллиметра до 15-20 см, а иногда и крупнее.

Чашевидный у пецицы
Часто плодовые тела образуются не на мицелии, а на его более или менее массивных сплетениях — стромах, имеющих различные размеры, окраску и консистенцию. На стволах и ветвях лиственных деревьев, особенно березы, можно встретить довольно крупные (до 4 см и более) полушаровидные или шаровидные углистые стромы дальдинии концентрической. На их разломе хорошо видны концентрические кольца. На поверхности стромы образуются перитеции, погруженные в нее по самый носик. После созревания и выбрасывания из них аскоспор нарастает новый слой стромы и в нем снова образуются перитеции. Этот процесс повторяется многократно и приводит к образованию слоистости.

В виде шляпки на ножке у леотии
Совсем иное строение и вид имеют стромы у спорыньи. Склероции этого гриба после перезимовки прорастают головчатыми стромами красноватого цвета с головкой диаметром 1 -1,5 мм. По периферии головки стромы развиваются перитеции. А у циттарий, обитающих в Южном полушарии на деревьях из рода нотофагус, образуются плотные группы крупных стром желтоватого или оранжевого цвета, на которых появляются многочисленные апотеции. Ч. Дарвин первый обратил внимание на эти интересные и необычные грибы, в массе встречающиеся в лесах Южной Америки, и описал их в своем «Путешествии натуралиста вокруг света». Он отметил также, что на Огненной Земле местные жители собирают зрелые стромы этих грибов и едят их сырыми. Стромы имеют сладковатый вкус, а по запаху напоминают шампиньоны.

Алеврия оранжевая
Для того чтобы увидеть базидии, нужно взять плодовое тело любого шляпочного гриба и отделить от него одну из пластинок или часть трубочек, расположенных на нижней поверхности шляпки. На поперечном срезе пластинки или трубочки под микроскопом виден, как и у дискомицетов, гимений, покрывающий поверхность пластинок или выстилающий полость трубочек. Однако у шляпочных грибов он выглядит иначе. Споры располагаются не внутри клеток, как у сумчатых грибов, а на поверхности булавовидных клеток, на выростах — стеригмах. На каждой такой клетке (базидии) образуется обычно по 4 базидиоспоры. Кроме базидий, в гимении можно видеть и другие элементы, например цистиды различной формы, выступающие над слоем базидий и защищающие его от соприкосновения с соседними пластинками и другими поверхностями.
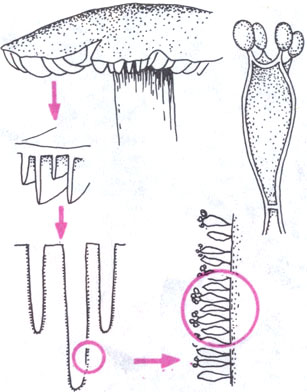
Базидии образуют плотный слой на пластинках шляпочного гриба
Базидии образуются у большой группы базидиальных грибов, или базидиемицетов, насчитывающей около 30 000 видов, однако не у всех они развиваются на сложно устроенных плодовых телах. У немногих из них базидии образуются непосредственно на мицелии (например, у экзобазидиума, дающего белые или розоватые пятна на деформированных листьях брусники). У головневых и ржавчинных грибов, паразитирующих на растениях, формируются покоящиеся споры, которые после перезимовки прорастают базидией. Плодовые тела того или иного строения имеются у большинства базидиомицетов, причем крайне разнообразные. Они могут иметь вид рыхлого прижатого налета или распростертых корочек разного цвета, встречающихся чаще всего на древесине, реже на почве и остатках травянистых растений. Гимений расположен у них на верхней стороне. Часто встречаются плодовые тела в виде палочек или булав, разветвленных кустиков, растущих на почве. На деревьях и гниющей древесине можно видеть плодовые тела копытообразные или в виде боковых черепитчато расположенных шляпок, коралловидно-разветвленные и т. д. Встречаются шляпочные грибы, плодовые тела которых состоят из шляпки и ножки.

Шляпочные грибы спороносят так обильно, что часто покрывают своими спорами окружающие их растения и почву
Все плодовые тела имеют разнообразную окраску и консистенцию, но при всем их разнообразии их объединяет одно — базидии образуют на всей поверхности плодового тела или части его (гименофоре) гимений, расположенный, как правило, открыто. Поэтому группа базидиомицетов с таким типом плодового тела получила название гименомицетов в отличие от гастеромицетов, у которых плодовые тела замкнуты до полового созревания базидио-спор. У последних базидии располагаются внутри плодового тела, там образуют базидиоспоры, а освобождение базидиоспор происходит уже после их отделения от базидий.
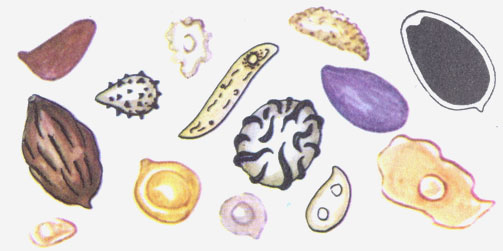
Споры шляпочных грибов разнообразны по форме и окраске
Эволюция плодовых тел гименомицетов шла, как и у других грибов, в направлении увеличения споровой продукции и совершенствования способов их распространения. Это особенно важно у гименомицетов, которые, как правило, не образуют митоспор, поэтому для расселения гриба служат базидиоспоры.
У многих базидиомицетов мицелий живет долго, часто много лет, и на нем каждый год или реже образуются плодовые тела с базидиоспорами, а у некоторых трутовиков и плодовые тела многолетние. Если одно плодовое тело красного мухомора за 5-6 дней своего существования образует до 575 млрд. спор, а на каждом мицелии ежегодно плодовых тел образуется несколько, можно представить, как велика споровая продукция одного мицелия мухомора за всю его жизнь.

Аурискальпиум, растущий на сосновых шишках
Увеличение числа спор, образующихся на одном плодовом теле, достигается у гименомицетов увеличением его поверхности, несущей гимений, — гименофора. У рогатиковых грибов с гладким гименофором происходит сильное расчленение и ветвление всего плодового тела, например у многочисленных рамарий, клавикороны, спарассиса — «грибной капусты» и других представителей этой группы.

Пластинцатый шляпочный гриб
У большинства же гименомицетов эволюция пошла по другому пути, пути совершенствования гименофора. Вместо гладкого гименофора у многих из них сформировался гименофор различного строения — складчатый (например, у настоящего домового гриба и у лисички), в виде разнообразных шипиков, гребенчатых пластин — у ежевиковых (ежевика желтого, герициума коралловидного и др.), трубочек разной длины и диаметра, внутренняя поверхность которых выстлана гимением, — у многочисленных трутовых и некоторых шляпочных грибов и, наконец, в виде пластинок, поверхность которых покрывает гимений, — у большинства шляпочных грибов. В результате увеличения гименофора многие гименомицеты образуют огромные количества спор.

Рогатики и экзобазидиум на листьях брусники

Рогатики и экзобазидиум на стеблях брусники
Если мы отрежем шляпку у шляпочного гриба, положим ее на белую или черную бумагу (в зависимости от окраски пластинок или трубочек) и покроем стеклянным колпаком или банкой, то уже через несколько часов на бумаге можно будет видеть массу спор гриба (споровый порошок), образующих рисунок, точно соответствующий расположению элементов гименофора. Споровый порошок часто можно увидеть и в природе, например на шляпках осенних опенков, расположенных в группах под другими плодовыми телами.
Тип гименофор базидиальных грибов (шиповидный)
При массовом освобождении базидиоспоры заносятся конвекционными токами воздуха на поверхность шляпки того же плодового тела. Это можно наблюдать, например, у сыроежек с красной или темной окраской шляпки или у плоского трутовика. Среднее плодовое тело навозника образует более миллиарда базидиоспор, а лишь на 1 см2 плодового тела настоящего домового гриба за сутки образуется до 35 млн. спор. Освобождение базидиоспор происходит путем их активного отбрасывания от базидии.

Тип гименофор базидиальных грибов (трубчатый)
В основе этого процесса, как и при выбрасывании аскоспор из сумок, лежит гидролиз гликогена и повышение осмотического давления внутри базидии. Через канал стеригмы давление передается базидиоспоре и она, получив толчок, отбрасывается на небольшое расстояние, обычно в доли миллиметра, а затем переносится токами воздуха.

Тип гименофор базидиальных грибов (пластинчатый)
В процессе эволюции плодовых тел у гименомицетов наблюдается перенос гименофора с верхней поверхности плодового тела (так он расположен на распростертых по субстрату плодовых телах) на его нижнюю поверхность (у плодовых тел, прикрепленных к субстрату боком, например у трутовиков, и шляпочных грибов). В этом случае после активного отбрасывания базидиоспоры начинается ее свободное падение, скорость которого зависит от массы споры. У шампиньона базидиоспора падает со скоростью 1 мм/с, а у других грибов — до 3 мм/с, Достаточно продолжительный период свободного падения базидиоспор значительно повышает вероятность переноса их токами воздуха. Однако в этом случае очень важной оказывается правильная, строго вертикальная ориентация трубочек или пластинок по отношению к поверхности земли, так как даже при небольшом их наклоне большая часть базидиоспор попадет на соседнюю пластинку или останется в трубочке. Но этого не происходит, так как плодовым телам гименомицетов свойствен геотропизм, обусловленный земным тяготением: их гименофор всегда ориентирован правильно.

Споровые отпечатки
Очень четко геотропизм проявляется у трутовиков. Их многолетние плодовые тела часто развиваются на дереве, стоящем вертикально, и их трубочки направлены вниз. После того, как дерево упадет, трубочки оказываются направленными к земле боком и споры в них задерживаются и не выпадают. Такие трубочки быстро зарастают стерильным мицелием, а гименофор начинает развиваться на той части плодового тела, которая обращена к земле, или начинает формироваться новое, правильно сориентированное по отношению к земле плодовое тело.

Если плодовое тело шляпочного гриба развивается на пне или стволе дерева, его ножка изгибается и пластинки распологаются строго вертикально
Совершенствование формы плодовых тел гименомицетов сопровождалось и дифференциацией гиф, из сплетения которых они состоят. Если наиболее примитивные плодовые тела, например у кортициевых, обитающих на влажной древесине, ветках, в лесной подстилке, где они образуют рыхлые или кожистые пленки или корочки, состоят из однородных тонкостенных и обильно ветвящихся гиф, то у трутовиков с копытообразными плодовыми телами они сложены гифами двух или трех типов: кроме тонкостенных, в них можно видеть толстостенные, малоразветвленные скелетные гифы и такие же толстостенные, но обильно ветвящиеся связывающие гифы, выполняющие механические функции. В плодовых телах млечников имеются специальные гифы, содержащие млечный сок, — млечные ходы.
Пожалуй, наиболее распространенный тип плодового тела у гименомицетов — это шляпочный гриб, т. е. плодовое тело, состоящее из шляпки на ножке. Плодовые тела этого типа обычно имеют мягкомясистую консистенцию и быстро загнивают, реже они бывают восковатой или кожистой консистенции. У негниючников они не загнивают, о чем говорит само их название, а высыхают в сухую погоду, вновь оживая после дождя. У навозников плодовое тело в зрелом возрасте расплывается (автолизируется). Шляпка и ножка могут быть самой разнообразной формы, причем их форма всегда постоянна и характерна для того или иного вида. Шляпка покрыта сверху кожицей, окраска и характер которой разнообразны. Окраска ее обычно определяет окраску шляпки, а кожица может быть гладкой, бархатистой, чешуйчатой и т. д., сухой или слизистой — она защищает мякоть шляпки от высыхания и повреждений. Мякоть шляпки может иметь различную консистенцию от плотной до рыхлой и губчатой, окраска ее обычно светлая, белая, желтоватая, светло-охряная или буроватая, однако на разломе или разрезе у некоторых грибов. например синяка, подосиновика, поддубника и др., ее окраска меняется: мякоть синеет, краснеет и темнеет. В мякоти млечников — рыжиков, груздей и др. — содержится бесцветный, белый или яркоокрашенный млечный сок, который на воздухе может изменять окраску.

Пример геопропизма у трутовиков. Плодовые тела всегда расположены на стволах деревьев так, что их трубочки направлены строго вертикально к поверхности земли
На нижней стороне шляпки расположен гименофор в виде пластинок или трубочек, очень редко в виде шипиков (у ежевика желтого) или складок (у лисичек). Пластинки по-разному прикрепляются к ножке: могут быть свободными, не сросшимися с ножкой, иногда они прикрепляются к ней на разном протяжении и бывают от низбегающих по ножке до выемчатых. Окраска пластинок и трубочек тоже разнообразна — от белой до темной, даже черной и обычно зависит от окраски базидиоспор, порошок которых может быть белым, желтоватым, розовым, охряным, оливковым, коричневым, фиолетово-бурым, пурпурно-бурым и черным.
После падения ствола дерева на старом плодовом теле под углом к нему выросло новое
Молодое плодовое тело часто окружено мицелиальным сплетением — общим покрывалом. По мере роста покрывало разрывается, а его остатки сохраняются у основания ножки в виде мешковидной или приросшей вольвы, а на шляпке — в виде разбросанных по ее поверхности лоскутов или хлопьев, например у мухомора. Иногда образуется покрывало другого типа, соединяющее край шляпки с верхней частью ножки и прикрывающее развивающийся гименофор, — его называют частным. Такое покрывало образуется у мухоморов, шампиньонов, некоторых маслят и других грибов и после разрыва сохраняется на ножке в виде кольца. У некоторых видов оно волокнистое или паутинистое (у паутинников), а также может быть слизистым (у мокрухи).
Гастеромицеты, как и гименомицеты, часто образуют очень большое количество базидиоспор. У дождевиков, например, их число достигает 7 биллионов, а в огромных плодовых телах лангермании гигантской их более 100 триллионов. У гастеромицетов не происходит активного отбрасывания базидиоспор от базидий, они освобождаются еще внутри оболочки плодового тела в результате разрушения базидий. Оболочка плодового тела (перидий) у гастеромицетов обычно хорошо развита и может состоять из одного или нескольких слоев. Ее разрушение и освобождение базидиоспор происходят разными способами.
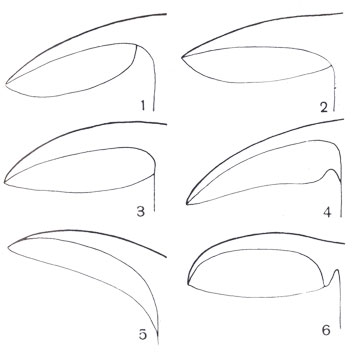
Типы расположения пластинок по отношению к ножке у шляпочных грибов: 1 — свободные; 2, 3 — приросшие; 4 — приросшие зубцом; 5 — нисходящие; 6 — приросшие к выросту ножки — коллариуму.
Как и у гименомицетов, у гастеромицетов эволюция шла по пути повышения эффективного рассеивания спор. У дождевиков и порховок плодовые тела вскрываются относительно небольшим отверстием на вершине, через которое при малейшем сотрясении, ударах капель дождя или порывах ветра вылетают базидиоспоры. Высыпание спор у этих грибов облегчается тем, что в их плодовых телах, в споровой массе, находятся нити капиллиция, при гигроскопических движениях которого масса разрыхляется. У головачей, или кальваций, перидий в верхней части плодового тела полностью разрушается, и оно приобретает форму, чаши. У близкой к ним лангермании гигантской перидий при созревании плодового тела растрескивается и отваливается неправильными кусками, обнажая споры. У звездовиков, называемых также земляными звездочками, наружный перидий состоит из трех слоев. Разрываясь на несколько остроконечных лопастей, он отгибается вниз в виде звездочки, за что эти грибы и получили свое название. У некоторых звездовиков он при этом еще и расслаивается, причем внутренний его слой выворачивается, поднимая плодовое тело, одетое внутренним перидием, над поверхностью почвы, как на ножках.
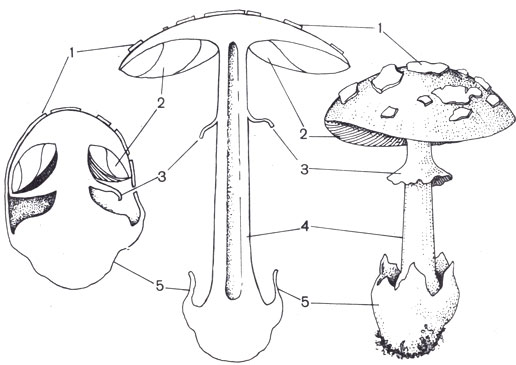
Схема строения плодового тела на разных стадиях развития: 1 — общее покрывало или его остатки в виде чешуек на шляпке; 2 — пластинки; 3 — частное покрывало и его остатки в виде кольца на ножке; 4 — ножка; 5 — вольва — остаток общего покрывала
Среди гастеромицетов есть и такие, у которых внутренняя часть плодового тела со спорами (глеба) может активно отбрасываться на большое расстояние — до нескольких метров. У сфероболюса звездчатого, маленькие, около 1-2 мм диаметром, шаровидные плодовые тела которого часто можно встретить на гниющей древесине, перидий многослойный, причем слои имеют разное строение. После созревания базидиоспор он разрывается от вершины к основанию звездообразно на 6-8 лопастей, расслаивается и внутренний слой быстро выворачивается наизнанку, отбрасывая глебу. У близких к сфероболюсу гнездовки и круцибулюма, или бокальчика, также обитающих на гнилой древесине, плодовые тела имеют вид крошечных чашечек или бокальчиков размером не более 1 -1,5 см, в которых лежат, как яйца в гнезде, округлые уплощенные перидиоли, содержащие внутри базидиоспоры и прикрепленные спирально закрученным канатиком к стенке чашечки. Если капля дождя упадет в такую чашечку, она разбрызгивается и выбрасывает из чашечки перидиолю. Канатик служит для прикрепления выброшенной перидиоли к субстрату — растениям, их остаткам и т. п.

Молодые дождевики

Зрелые дождевики
Большая группа гастеромицетов — фаллюсовые — адаптировалась к распространению базидиоспор при помощи насекомых. Глеба у них после созревания расплывается в слизь, содержащую базидиоспоры. Слизь имеет очень сильный запах падали, привлекающий мух и других насекомых, которые и переносят базидиоспоры. У часто встречающейся у нас в лесах веселки обыкновенной темно-зеленая слизистая глеба располагается, как шляпка, на длинной губчатой ножке — рецептакуле. У тропических грибов — решеточника и цветохвостника — рецептакул более сложного строения: у первого он имеет вид сетчатого яйцевидного образования, внутри которого располагается слизистая глеба, а у второго рецептакул состоит из короткой ножки, от которой отходят длинные лопасти, соединенные на вершине или звездчато отгибающиеся. Рецептакул у этих грибов ярко окрашен, что также привлекает к ним насекомых.

Звездовики, которых называют ещё земляными звездочками за необычную для грибов форму
Развитие и размножение высших грибов
Как и все живое на Земле, гриб начинает свою жизнь с одноклеточного состояния — споры. Размеры спор грибов не превышают 10-25 мк в поперечнике.
Внешний вид их поразительно разнообразен. Достаточно сказать, что внешний вид спор служит одним из важных систематических признаков классификации грибов.
Спора прорастает, путем последовательного митотического деления разрастается в длину, образуя тонкие нити-гифы.
Гифы, вырастающие из одной споры, расходятся от нее во все стороны лучами, ветвятся и образуют первичный мицелий.
Дальнейшее развитие однополого первичного мицелия шляпочных базидиальных грибов включает своеобразную форму полового процесса. При соприкосновении кончиков гиф двух разнополых мицелиев происходит разрушение оболочек и слияние протопластов. Ядра сливающихся клеток не соединяются, и образуется двухъядерная клетка — дикарион. Этот процесс именуется плазмогамией. При дальнейшем делении двухъядерных клеток возникает своеобразный феномен, обеспечивающий каждой клетке получение и мужского и женского ядра. В начале деления на одном — заднем — конце клетки гифы образуется отросток — «крючок», или «пряжка», и переднее ядро спускается в этот отросток, после чего каждое из ядер делится на два. Одно из дочерних ядер переднего ядра из «крючка» отходит вперед, в переднюю клетку, где остается и одно из дочерних ядер заднего ядра после образования перегородки. При этом дочерние ядра заднего и переднего ядра вместе с «крючком» остаются в задней клетке. Так образуются две клетки, каждая с двумя разнополыми ядрами.
Из этих клеток путем последовательного деления разрастается вторичный двухъядерный мицелий. Он может при благоприятных внешних условиях образовать наземное плодовое тело — гриб. Плодовое тело представляет собой компактное сплетение гиф и служит органом для образования спор.
Гифы грибного мицелия могут, особенно у дереворазрушающих грибов, соединяться в толстые образования — шнуры-ризоморфы, которые в почве или на ином субстрате тянутся очень далеко. Иногда гифы образуют компактные темные образования — склероции. В этих и других разновидностях мицелия гифы никогда не сливаются, идут тесно и параллельно, как и в плодовом теле.
Споры у базидиальных шляпочных грибов образуются на нижней стороне шляпки. Здесь под мякотью шляпки вырастает гименофор (несущий гимений), состоящий из пластинок или трубочек, иногда просто складок, спускающихся на ножку, или шипиков и сосочков. На гименофоре развивается спорообразующий слой — гимений. В нем гифы заканчиваются и образуют концевые удлиненные двухъядерные клетки своеобразного строения — в форме цилиндра, булавы и др., — именуемые базидиями. Между плодоносящими базидиями располагаются тоже удлиненные клетки гиф — цистиды и парафизы. Эти одноклеточные или многоклеточные образования в форме мешочков бесплодны и выполняют, вероятно, питательную функцию в отношении базидий.
Процесс образования базидиоспор начинается со слияния обоих ядер базидий в одно большое, что трактуется как половой процесс — кариогамия. Путем быстрых последовательных делений большое ядро превращается в четыре дочерних, отделяющихся друг от друга перегородками. Далее каждое из новых ядер проникает в один из концевых отростков базидии — через стеригму, на конце которой происходит в течение нескольких часов созревание спор.
Плодовое тело, гименофор, гимений и базидии следует рассматривать как производные мицелия.
Описанный выше процесс размножения свойствен почти всем базидиальным грибам, но встречаются и отклонения от него. Так, у гриба Coprinus sp. первичный мицелий путем слияния своих же клеток становится двухъядерным, способным к развитию плодовых тел. У некоторых видов клетки мицелия могут содержать одно двуполое ядро, у других не образуется «пряжка». Образование «пряжки» характерно для размножения многих высших базидиальных грибов.
Для сравнения кратко упомянем о способах размножения других групп грибов. Споры образуются уже у слизевиков, которых раньше относили к классу низших грибов, а теперь выделяют в самостоятельный тип. Обычно слизевики существуют в виде одноядерных гаплоидных голых амеб, питающихся бактериями и способных к размножению. В определенный момент все амебы сливаются в один большой многоядерный плазмодий, в котором образуются споры, представляющие собой не что иное, как те же инцистированные амебы. При прорастании спор из них вытекают амебы. Цикл развития истинных слизевиков — миксомицетов, а также хитридиомицетов, оомицетов и, наконец, аскомицетов (сумчатых грибов) довольно сложен и включает в себя как фазу бесполого, так и фазу полового размножения. У многих из них образуются половые органы — оогонии и антеридии.
Грибы отличаются своим необычайно обильным плодоношением. Так, плодовое тело шампиньона — Agaricus campestris величиной в 8 см в поперечнике образует 1800 млн спор, выдавая каждую секунду по 40 млн спор. На 214 пластинках гриба Coprinus sp. образуется до 5000 млн спор, по 100 млн в час, а еще больше — до 1 000 млн спор имеется у ежовика — Sarcodon imbricatus.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.